Em 1835, durante a viagem do HMS Beagle, Charles Darwin coletou nas ilhas Galápagos pequenas aves que lhe pareceram variações de um mesmo tipo. Os tentilhões de Darwin, como foram reconhecidos posteriormente, de fato constituem um conjunto de 14 espécies intimamente aparentadas distribuídas entre as ilhas de Galápagos e na ilha de Cocos. Essas espécies consideradas um dos casos mais emblemáticos de radiação adaptativa, Essas apresentam grande diversidade no formato do bico que está diretamente associada aos diferentes nichos ecológicos explorados, como sementes duras, insetos e flores de cactos[3,6].Essas diferenças externas refletem alterações no esqueleto craniofacial e já estão presentes no momento da eclosão. Ou seja, não são moldadas pela experiência individual, mas estabelecidas durante o desenvolvimento embrionário. O formato do bico é geneticamente determinado e produzido antes mesmo de o animal interagir com o ambiente[3,4].
Sob a perspectiva da biologia evolutiva do desenvolvimento (evolutionary developmental biology, Evo-Devo), os tentilhões oferecem um modelo privilegiado para compreender como a radiação adaptativa emerge da modulação de programas de desenvolvimento. A diversificação rápida do grupo nas ilhas decorrente da ocupação de distintos nichos ecológicos e envolveu também episódios de hibridação introgressiva, nos quais o fluxo gênico entre linhagens contribuiu para recombinar variantes já existentes[6,7]. Assim, a diversidade morfológica observada resulta da interação entre seleção natural, plasticidade dos sistemas de desenvolvimento e intercâmbio genético, evidenciando que a evolução pode explorar variações reguladoras preexistentes e redistribuí-las entre populações em processo de divergência[3,4].
Assim, o bico dos tentilhões deixou de ser apenas um exemplo didático de radiação ecológica e tornou-se uma janela privilegiada para compreender como a evolução opera em termos moleculares. A seleção natural não cria estruturas do zero; ela atua sobre programas de desenvolvimento preexistentes, ajustando qualitativa e quantitativamente seus parâmetros reguladores.
 Figura 1. (a) Ilhas Galápagos, como a Ilha Floreana, são ilhas vulcânicas visitadas por Charles Darwin em 1835; (b) arbustos de cactos figo-da-índia (Opuntia helleri) na Ilha Genovesa (Ilha Tower); (c) flores do geiger-amarelo (Cordia lutea); (d) macho do tentilhão-terrestre-grande (Geospiza magnirostris) cantando durante a estação chuvosa; (e) fêmea do tentilhão-terrestre-grande (G. magnirostris) na Ilha Genovesa; (f) fêmea do tentilhão-terrestre-médio (G. fortis) na Ilha Santa Cruz; (g) macho do tentilhão-de-cacto-grande (G. conirostris); (h) macho do tentilhão-de-bico-afiado (G. difficilis) alimentando-se de flores de cacto na Ilha Genovesa; (i) macho do tentilhão-dos-bosques (Certhidea fusca) cantando ao lado de seu ninho. Fonte: Abzhanov, A. (2010).
Figura 1. (a) Ilhas Galápagos, como a Ilha Floreana, são ilhas vulcânicas visitadas por Charles Darwin em 1835; (b) arbustos de cactos figo-da-índia (Opuntia helleri) na Ilha Genovesa (Ilha Tower); (c) flores do geiger-amarelo (Cordia lutea); (d) macho do tentilhão-terrestre-grande (Geospiza magnirostris) cantando durante a estação chuvosa; (e) fêmea do tentilhão-terrestre-grande (G. magnirostris) na Ilha Genovesa; (f) fêmea do tentilhão-terrestre-médio (G. fortis) na Ilha Santa Cruz; (g) macho do tentilhão-de-cacto-grande (G. conirostris); (h) macho do tentilhão-de-bico-afiado (G. difficilis) alimentando-se de flores de cacto na Ilha Genovesa; (i) macho do tentilhão-dos-bosques (Certhidea fusca) cantando ao lado de seu ninho. Fonte: Abzhanov, A. (2010).A crista neural: onde a forma começa
Para entender como a diversidade dos bicos surgiu, pesquisas de Biologia do Desenvolvimento investigaram uma população de células muito peculiares, as células da crista neural craniana (CCNs), geradas nas etapas iniciais do desenvolvimento embrionário. As células da crista neural, encontradas apenas no desenvolvimento dos vertebrados, surgem na borda do tubo neural em desenvolvimento e migram para todas as regiões do embrião, incluindo células na região da face que se tornam a principal fonte do mesênquima facial, o tecido embrionário que dará origem a quase todo o esqueleto e outros tecidos conjuntivos do bico[9]. Os embriões de aves foram os primeiros modelos usados para mapear o destino dessas células, no caso das aves, revelando que as partes superior e inferior dos bicos, além de grande parte do crânio, são formadas a partir delas.
O primeiro estudo, realizado em 2003, a fazer a descoberta que as CCNs funcionam como “arquitetas” da forma do bico, realizou um experimento no qual houve a troca de CCNs entre embriões de codorna e de pato[8]. Os resultados foram surpreendentes: embriões de pato desenvolveram bicos curtos e pontiagudos (típicos de codorna), enquanto embriões de codorna desenvolveram bicos longos e largos (típicos de pato). Essa descoberta evidenciou que a crista neural não é apenas um material de construção passivo, mas sim o centro de comando da morfogênese facial[8]. As células da crista neural carregam programas moleculares intrínsecos e específicos de cada espécie que determinam o tamanho e o formato do bico[4].
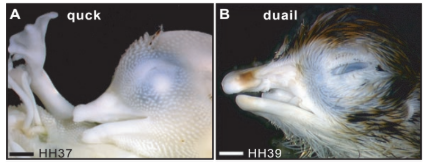
Além disso, o que tornou tal experimento ainda mais revelador, foi observar que as CCNs foram capazes de reprogramar os tecidos do embrião hospedeiro, instruindo tecidos adjacentes a seguir o plano de desenvolvimento da espécie doadora. Mais do que terem fornecido instruções para todo o tecido ao seu redor, essas células também mantiveram o próprio cronograma de maturação celular característico da espécie doadora[9].
BMP4: tornando o bico mais largo e profundo
Entre os genes investigados nos estudos comparativos de desenvolvimento craniofacial, destacou-se BMP4 (Bone Morphogenetic Protein 4), membro da família TGF-β, amplamente reconhecida por sua atuação na regulação da proliferação, diferenciação e sobrevivência celular durante a embriogênese. No contexto craniofacial, BMP4 exerce papel determinante na expansão e padronização do mesênquima derivado da crista neural craniana, tecido embrionário que dará origem ao bico[1].
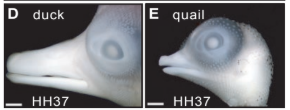
Observou-se que espécies de bico robusto, como Geospiza magnirostris, apresentam expressão mais precoce, mais intensa e espacialmente expandida de BMP4 no mesênquima do processo frontonasal. Em termos mecanísticos, isso significa que o sinal molecular responsável por estimular a proliferação de progenitores osteocondrogênicos é ativado antecipadamente e com maior magnitude. Como consequência, ocorre aumento na taxa proliferativa dessas populações celulares, promovendo expansão concomitante da largura e da profundidade do bico[1].
A relação observacional foi submetida a teste funcional utilizando embriões de Gallus gallus como modelo experimental. Por meio do vetor retroviral RCAS, induziu-se a superexpressão de BMP4 especificamente na região do processo frontonasal. Os embriões apresentaram aumento significativo nas dimensões laterais e dorsoventrais do bico, com expansão proporcional da estrutura óssea interna que sustenta o bico subjacente[1].
Esses dados sustentam a interpretação de que a robustez do bico não decorre do surgimento de um novo gene, mas de uma modulação quantitativa, temporal e espacial na expressão de um gene já integrante do repertório genômico[1].
Calmodulina: alongando o bico
Se BMP4 explica a expansão em largura e profundidade, permanecia a questão acerca da origem dos bicos longos e pontiagudos, adaptados à exploração de flores e cactos.
Análises de expressão gênica em larga escala identificaram um candidato inesperado: calmodulina (CaM), proteína ligante de cálcio (Ca²⁺) que atua como mediadora central em múltiplas vias de sinalização intracelular, incluindo a ativação da quinase dependente de calmodulina, CaMKII. Até então, sua participação direta na morfogênese craniofacial não havia sido claramente estabelecida[2].
Espécies de bico alongado, como Geospiza conirostris, apresentaram níveis elevados de expressão de CaM na região distal do bico em desenvolvimento. A hipótese funcional foi novamente testada em embriões de Gallus gallus, por meio da ativação aumentada da via CaM/CaMKII[2].Observou-se um aumento significativo no comprimento do bico, sem alterações relevantes na largura ou na profundidade. Diferentemente de BMP4, cuja ação promove expansão bidimensional, a via CaM/CaMKII atua predominantemente ao longo do eixo anteroposterior, modulando o alongamento distal da estrutura[2].
Um princípio central: modularidade
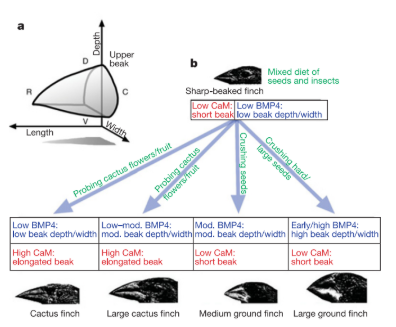
Conjuntamente, esses resultados revelaram um princípio fundamental da morfogênese do bico: sua organização é modular. Nesse processo , BMP4 atua predominantemente na determinação da largura e da profundidade da estrutura, ao passo que a via CaM/CaMKII regula principalmente o eixo anteroposterior, controlando o alongamento do bico.
Sendo assim, os diferentes eixos de crescimento podem, portanto, variar de maneira relativamente independente. Essa independência funcional aumenta a chamada evolucionabilidade da estrutura — isto é, sua capacidade de gerar variações morfológicas adaptativas sem comprometer a integridade do desenvolvimento[2].Assim, os estudos sobre BMP4 e a via CaM/CaMKII revelam que o bico dos tentilhões é construído a partir de módulos de crescimento relativamente independentes. Enquanto BMP4 atua principalmente na expansão lateral e dorsoventral da estrutura, a via da calmodulina regula o alongamento ao longo do eixo anteroposterior. Essa organização modular permite que diferentes dimensões do bico variem de maneira parcialmente independente durante o desenvolvimento, criando múltiplas combinações morfológicas possíveis a partir de um mesmo programa básico de desenvolvimento[2].
Abreviações: C, caudal; D, dorsal; R, rostral; V, ventral. Fonte: Abzhanov, A. et al. (2006).
Genômica, ALX1 e evolução em tempo real
O gene ALX1 (ALX Homeobox 1) codifica um fator de transcrição com um homeodomínio tipo-paired (paired-type homeodomain) que é fundamental para o desenvolvimento craniofacial em vertebrados. Ele atua na regulação de estruturas derivadas do mesênquima facial e do primeiro arco branquial, além de influenciar diretamente a migração das células da crista neural craniana[7]. A importância biológica deste gene é evidenciada pelo fato de que, em humanos, mutações que causam a perda de função do ALX1 resultam em displasia frontonasal, uma condição que gera malformações faciais graves, como fissuras nos tecidos do rosto e redução severa do tamanho dos globos oculares[5].
A descoberta do papel central do ALX1 na evolução dos tentilhões de Darwin foi possibilitada, em 2015, pelo sequenciamento de genoma completo de 120 indivíduos de todas as espécies conhecidas do grupo[7]. Através de uma varredura genômica, os cientistas identificaram uma região de 240 quilobases que apresentava a diferenciação mais forte entre espécies com bicos robustos (blunt) e pontiagudos (pointed). Esta região continha o gene ALX1, tornando-o o principal candidato molecular para explicar a diversidade morfológica do formato do bico[7].
Existem dois haplótipos (conjuntos de variantes genéticas) principais para este gene: o haplótipo B (robusto) e o haplótipo P (pontiagudo)[7]. Entre eles, foram identificados 335 SNPs (Single Nucleotide Polymorphisms) fixos, incluindo duas mutações missense (L112P e I208V) que alteram aminoácidos altamente conservados na proteína. Essas mutações são classificadas como “danosas” à função original da proteína, sugerindo que elas foram fundamentais para a transição evolutiva entre os diferentes formatos de bico, afetando tanto a sequência codificante quanto as regiões reguladoras do gene[7].
Na prática, essa diferenciação molecular revela que o formato pontiagudo (P) é a condição ancestral, enquanto o formato robusto (B) surgiu a partir do acúmulo dessas variantes genéticas derivadas[7]. As duas mutações missense específicas são encontradas justamente no haplótipo B, sugerindo que a alteração funcional na proteína ALX1 foi um dos gatilhos moleculares que permitiu a transição de bicos finos para formatos robustos que são observados nos tentilhões terrestres. Dessa forma, o bico é ativamente remodelado por uma proteína cuja função original foi modificada por essas mutações, criando uma associação direta entre o genótipo e a anatomia dos bicos das aves[7].
A variação no ALX1 não ocorre apenas entre espécies diferentes, mas também é responsável pela diversidade dentro de uma única espécie, como o tentilhão terrestre médio (Geospiza fortis). A análise dos genomas sequenciados em 2015 demonstrou que aves homozigotas para o alelo P possuem bicos visivelmente mais pontiagudos, enquanto homozigotos B possuem bicos mais robustos e profundos, com os heterozigotos exibindo formas intermediárias. O haplótipo B, associado a bicos robustos, é uma variante antiga que surgiu há centenas de milhares de anos, precedendo a radiação de muitos tentilhões terrestres e arbóreos[7].
O monitoramento genômico de longo prazo na ilha Daphne Major, abrangendo cerca de 30 anos, descrito em um estudo de 2023, permitiu observar a evolução em tempo real relacionada a este gene. Pesquisadores descobriram que a frequência dos alelos do ALX1 (localizado no locus genético G07) oscila rapidamente em resposta a eventos climáticos e pressões seletivas, como secas severas que alteram a disponibilidade de alimentos. Em G. fortis, o locus do ALX1 explica aproximadamente 14% da variação herdável no formato do bico, demonstrando como variantes de grande efeito genético permitem adaptações rápidas em populações selvagens[5].
Além da seleção natural, a hibridação introgressiva desempenha um papel crucial na dinâmica do ALX1, permitindo que variantes genéticas “saltem” entre espécies. Por exemplo, tentilhões de bicos pontiagudos que adquiriram alelos para bicos robustos através de eventos de hibridação interespecífica puderam explorar novos nichos alimentares[5]. Evidências sugerem que o ALX1 pode atuar como parte de um supergene, um cluster de genes proximamente ligados que inclui o vizinho LRRIQ1, o qual possui domínios de ligação à calmodulina e pode interagir com outras vias de sinalização para coordenar transformações morfológicas complexas[5].
Qual a importância disso para a Biologia moderna?
A trajetória científica iniciada em 1835 com Charles Darwin, a partir de suas observações nas Ilhas Galápagos, evoluiu ao longo de quase dois séculos para um dos modelos experimentais mais complexos da biologia moderna[3]. Em conjunto, as evidências oriundas da embriologia experimental, da biologia molecular, da genômica populacional e do monitoramento de campo convergem para uma conclusão central da Biologia do Desenvolvimento moderna: a evolução morfológica é, em grande medida, evolução das redes regulatórias do desenvolvimento[9]. O enigma que começou com observações naturalistas nas ilhas Galápagos revela-se hoje como um exemplo paradigmático de como redes regulatórias, populações celulares embrionárias e variantes genéticas interagem para gerar diversidade adaptativa[3].
A crista neural craniana emerge como um ponto nodal onde informação genética é traduzida em arquitetura anatômica. Genes como BMP4, calmodulina e ALX1 não “criam” novas estruturas, mas modulam parâmetros de crescimento, proliferação, migração e diferenciação celular. A forma final do bico, portanto, não é produto de um único fator, mas da dinâmica integrada de redes de sinalização e fatores de transcrição atuando em tecidos embrionários específicos[2].
A Biologia do Desenvolvimento contemporânea amplia essa compreensão ao integrar diferentes escalas: do SNP ao organismo, do embrião à população. O caso de ALX1 demonstra que variantes de grande efeito podem ser mantidas, recombinadas por hibridação e rapidamente moduladas pela seleção natural em resposta a mudanças ambientais. Assim, a evolução deixa de ser vista apenas como um processo histórico distante e passa a ser observada como um fenômeno mensurável em tempo real, ancorado em mecanismos celulares e moleculares bem caracterizados[5]. A importância desse tema para a Biologia do Desenvolvimento moderna reside justamente nessa integração conceitual. Compreender como variações regulatórias se traduzem em diferenças morfológicas permite conectar genética, embriologia, ecologia e evolução em um único arcabouço explicativo. O bico dos tentilhões deixa, assim, de ser apenas um símbolo histórico da adaptação e torna-se um exemplo robusto de como a diversidade biológica emerge da modulação precisa e dinâmica dos programas que constroem os organismos desde seus primeiros estágios embrionários[4].
Referências
[1] ABZHANOV, Arhat et al. Bmp4 and morphological variation of beaks in Darwin’s finches. Science, v. 305, n. 5689, p. 1462-1465, 2004.
[2] ABZHANOV, Arhat et al. The calmodulin pathway and evolution of elongated beak morphology in Darwin’s finches. Nature, v. 442, n. 7102, p. 563-567, 2006.
[3] ABZHANOV, Arhat. Darwin’s Galapagos finches in modern biology. Philosophical Transactions of the Royal Society B: Biological Sciences, v. 365, n. 1543, p. 1001-1007, 2010.
[4] ABRAMYAN, John; RICHMAN, Joy M. Craniofacial development: discoveries made in the chicken embryo. International Journal of Developmental Biology, v. 62, n. 1-2-3, p. 97-107, 2018.
[5] ENBODY, Erik D. et al. Community-wide genome sequencing reveals 30 years of Darwin’s finch evolution. Science, v. 381, n. 6218, p. 1-9, 2023.
[6] GRANT, Peter R.; GRANT, Rosemary B. Darwin’s finches. Current Biology, v. 15, n. 16, p. 614-615, 2005.
[7] LAMICHHANEY, Sangeet et al. Evolution of Darin’s finches and their beaks revealed by genome sequencing. Nature, v. 518, p. 371-375, 2015.
[8] SCHNEIDER, Richard A.; HELMS, Jessye A. The Cellular and Molecular Origins of Beak Morphology. Science, v. 299, n. 5606, p. 565-568, 2003.9. SCHNEIDER, Richard A. How to tweak a beak: molecular techniques for studying the evolution of size and shape in Darwin’s finches and other birds. BioEssays, v. 29, p. 1-6, 2006.